الكروموسوم البشري 15q11-13 هو مكان معقد يحتوي على جينات مطبوع، فضلا عن مجموعة من ثلاثة GABA A مستقبلات الوحيدات (جبر) GABRB3 genes-، GABRA5 وGABRG3. حذف أو ازدواجية 15q11-13 الجينات جبر يحدث في العديد من الاضطرابات العصبية النمائية الإنسان بما في ذلك متلازمة برادر ويلي (PWS)، متلازمة أنجلمان (AS) والتوحد. يتم تخفيض GABRB3 البروتين التعبير أيضا في متلازمة ريت (RTT)، التي تسببها طفرات في MECP2 على Xq28. على الرغم من Gabrb3 وأعرب عن biallelically في مخ الفأر، توجد بيانات متضاربة بشأن الوضع يطبع من الجينات 15q11-13 جبر في البشر. باستخدام الترميز الأشكال النووية المنفردة وتبين لنا أن كل الجينات جبر ثلاثة يتم التعبير عن biallelically في 21 عينة الدماغ السيطرة، مما يدل على أن هذه الجينات لا مطبوع في القشرة الإنسان العادي. ومن المثير للاهتمام، وأربعة من ثمانية التوحد واحدة من خمس عينات الدماغ RTT أظهرت monoallelic أو منحرفا للغاية التعبير أليلية واحد أو أكثر جبر الجيني، مما يشير إلى أن التقلبات جينية من هذه الجينات هو شائع لظهور هذه الاضطرابات السلوكية. كشف الكمي في الوقت الحقيقي تحليل RT-PCR من PWS وAS العينات مع الأب والحذف 15q11-13 الأمهات تحيز التعبير الأب من GABRB3، في حين أظهرت عينات الدماغ RTT انخفاض كبير في GABRB3 وUBE3A. أظهر مناعي لونين وتسلسل بيسلفيت في الخلايا العصبية SH-SY5Y أن MeCP2 بربط المواقع الدليل السياسي الشامل مثيلة داخل GABRB3. أظهرت دراساتنا السابقة أن الاقتران مثلي 15q11-13 في الخلايا العصبية يتوقف على MeCP2 وعطلت في RTT والتوحد القشرة. جنبا إلى جنب، وتشير هذه النتائج إلى أن MeCP2 بمثابة منظم لونين للتعبير الأمثل لكل من الأليلات من GABRB3 في الخلايا العصبية.
الكروموسوم البشري 15q11-13 هو مكان معقد يحتوي على جينات مطبوع تشارك في اثنين من العصبية متلازمة الاضطرابات برادر ويلي (PWS) ومتلازمة أنجلمان (AS). PWS، وهو اضطراب يتميز فرط الأكل مما يؤدي إلى السمنة مع خفيفة الى معتدلة التخلف العقلي، وينتج عن نقص الأب من 15q11-13. AS، تتميز التخلف العقلي الشديد مع عدم وجود الكلام والضحك غير مناسب المفرط، والناجمة عن الأمهات 15q11-13 نقص
(1). غالبية PWS وAS الحالات (~70٪) نتيجة من
دي نوفو الحذف تمتد المنطقة 4 ميجا بايت من 15q11-13. disomy أحد الأبوين (UPD) من كروموسوم الأمهات سبب آخر شائع (~25٪ من الحالات) من PWS، في حين أن الأب UPD هو سبب نادر من AS. الطفرات داخل أعربت أمهات مطبوع
UBE3A الجينات، كافية للتسبب AS
(2، 3)، وتحدث في ~10٪ من الحالات
(1).
التوحد هو اضطراب النمو العصبي المعقدة التي تتميز ضعف في التفاعل الاجتماعي، والسلوكيات المحظورة والنمطية، والعجز في اللغة والتواصل
(4). بالرغم من وجود عنصر وراثي قوي لمرض التوحد، تم تحديد عدد قليل من الجينات المرشحة
(5). كروموسوم 15q11-13 هو واحد من المواضع الجينية المتورطين في التوحد، والازدواجية الأمهات في هذه المنطقة لا تزال واحدة من تشوهات الوراثية الخلوية الأكثر شيوعا وجدت في حالات التوحد مجهول السبب
(6). أدلة إضافية من الربط وجمعية دراسات متعددة تشير تشارك 15q11-13 الجينات في مرض التوحد
(+7 -
10).
متلازمة ريت (RTT) هو اضطراب النمو العصبي الشديد الذي يؤثر في المقام الأول الإناث والتي تسببها طفرات في العاشر مرتبطة
MECP2 الجيني، والميثيل الدليل السياسي الشامل بروتين 2
(11). يتميز RTT في أوائل التنمية ما بعد الولادة الطبيعية تليها خسارة مفاجئة من المعالم التنموية حول 6-18 شهرا من العمر مما يؤدي إلى غياب الكلام، والتخلف العقلي الشديد وفقدان حركات اليد هادفة
(12). مثل التوحد، يتم تصنيفها RTT كاضطراب النمو المتفشي (PDD)، وهو الوحيد من خمسة ثائق تصميم المشاريع مع سبب وراثي معروف
(4). في دراسة جزيئية الأخيرة من الدماغ البشري بعد الوفاة، سواء RTT وعينات التوحد أظهرت عيوب التعبير في اثنين من الجينات 15q11-13،
UBE3A وGABRB3، مما يشير إلى أن مسارات متداخلة وdysregulated في هذه الاضطرابات
(13).
كروموسوم 15q11-13 يحتوي على مجموعة من ثلاثة GABA A مستقبلات الجينات الوحيدات،
GABRB3، GABRA5 وGABRG3، ترميز مفارز مستقبلات الناقل العصبي المثبط للالغاما غاما حمض (GABA). وorthologues الماوس، الواردة ضمن كتلة syntenic على الصبغي 7qB5، يتم التعبير عن biallelically في الدماغ
(+14 -
+16)، ومع ذلك، توجد بيانات متضاربة حول الوضع يطبع من البشر 15q11-13 GABA A جينات مستقبلة الوحيدات
(17 -
21). اقترحت دراسة مبكرة من
GABRB3 التعبير في الخلد عداري الشكل والأنسجة البشرية الزغب العادية تفضيلية التعبير الجيني الأم
(20). جبريل
وآخرون. (21) الكشف في وقت لاحق 15q11-13 GABA تعبير مستقبلات في خطوط هجين بين الإنسان والفأر خلية جسدية متعددة تحتوي على الأم أو الأب الكروموسوم البشري 15، مما يشير إلى أن هذه الجينات هي غير مطبوع. ميغورو
آخرون. (17) يقوم بنقل الخلية بوساطة microcell لخلق الماوس الهجينة A9 إيواء إما العادي الأب أو الأم الكروموسوم البشري 15. إن الإنسان أعربت أبويا الجينات مطبوع احتفظت يطبع طبيعية في الهجينة A9، ومع ذلك، فإن GABA وأعرب عن جينات مستقبلة فقط من كروموسوم الأب. Bittel
وآخرون. (18، 19) أثبتت أدلة تدعم الأب التحيز التعبير عن GABA الإنسان جينات مستقبلة مع ميكروأري تحليلات. كلتا الدراستين، مقارنة الحذف وUPD خطوط الخلايا lymphoblastoid من PWS وAS المرضى، وجد التحيز الأب ثابت من
GABRB3 والتعبير
GABRA5 الجينات.
أدلة توحي إضافي ليطبع من الإنسان 15q11-13 GABA A مستقبلات يأتي من الخلافات توقيت تكرار أليل معين من تسلسل الجينوم بالقرب من GABA A جينات مستقبلة
(22)، وفروق في التوقيت النسخ المتماثل بين المتماثلات الأبوية هي سمة من الجينات مطبوع
(23) . كلاهما مطلوب مساهمات الكروموسومات الأبوية لتوقيت تكرار السليم لل15q11-13 GABA A جينات مستقبلة، مما يشير إلى التفاعلات
العابرة بين homologs الوالدين
(24). وعلاوة على ذلك، جمعية مثلي من 15q11-13 المجالات يحدث في الخلايا الليمفاوية الطبيعية، ولكن ناقصة في PWS وAS العينات، مما يدل على أن التفاعل
عبر ينطوي على جمعية المتماثلات مطبوع معاكس
(25). جمعية مثلي من 15q11-13 الأليلات
GABRB3 يحدث في الخلايا العصبية الناضجة داخل الدماغ، وناقصة في RTT والتوحد
(26).
وبما أن تورط 15q11-13 GABA A مستقبلات الجينات في الإنسان اضطرابات متعددة، بما في ذلك مرض التوحد، وتحديد نهائيا الوضع يطبع من هذه الجينات في الدماغ البشري أمر بالغ الأهمية لفهم دورها المحتمل في التسبب في اضطرابات النمو العصبي. في هذه الدراسة، تم استخدام الأشكال النووية المنفردة (النيوكلوتايد) في المنطقة ترميز
GABRB3، GABRA5 وGABRG3 للتحقيق في التعبير أليلية النصوص من القشرة الدماغية. منذ عدة جينات مطبوع، بما في ذلك
UBE3A، يحمل الخلايا العصبية محددة يطبع
(16، 27 -
+29)، وقد أجريت هذه الدراسة يطبع باستخدام ما بعد الوفاة أنسجة المخ البشري. والإنسان 15q11-13 GABA تم تحديد مستقبلات الجينات الوحيدات أن يكون التعبير biallelic على قدم المساواة في جميع العينات السيطرة، مما يدل على أن هذه الجينات لا مطبوع في العادية القشرة الأمامية. في المقابل، كشف تحليل التعبير أليلية أن أربع عينات التوحد مجهول السبب وعينة واحدة RTT أظهرت monoallelic أو منحرفا للغاية التعبير أليلية من GABA واحد أو أكثر من الجين المستقبل. وعلاوة على ذلك، أظهر التحليل الكمي
للGABRB3 نص في PWS وAS عينات الدماغ دليل على وجود تحيز التعبير الأب عندما متحدر من والدين 15q11-13 مساهمة تفتقر. هذه النتائج تفسر الملاحظات السابقة بالانحياز الأب من
GABRB3 التعبير، وتشير إلى أن تشوهات جينية في 15q11-13 التي تعمل على dysregulate
GABRB3 التعبير قد يكون أمرا شائعا نسبيا في الأفراد المصابين بالتوحد دون التعديلات الوراثية الخلوية يمكن اكتشافها.
عادة يتم التعبير عن الجينات 15q11-13 GABA A مستقبلات biallelically في القشرة، ولكن monoallelically أعرب في بعض العينات RTT والتوحد
من أجل تحديد بشكل قاطع إذا 15q11-13 GABA تعبير جينات مستقبلات المعرض مطبوع، تم تحليل التعبير أليلية في عينات الدماغ السيطرة من القشرة الأمامية، أدرجت منطقة برودمان 9. RTT والتوحد عينات الدماغ أيضا في تحليل لاختبار دليل على جينية التقلبات. تم عزل DNA و RNA من المجمدة عينات الدماغ بعد الوفاة مع فاصل زمني بعد الوفاة (PMI) <30 ساعة (التفاصيل في المواد التكميلية، الجدول S1). تم تحليل نسخة من كروموسوم رقم 15q11-13 من قبل مضان DNA
الموقع التهجين (FISH DNA)
في استخدام مسبار
GABRB3 (26)، تهجين لعينات الدماغ على ميكروأري الأنسجة
(30). وتم التحقق العادية disomic 15q11-13 مساهمات في جميع السيطرة، RTT وAUT عينات الدماغ (لا تظهر البيانات).
لتحديد الأشكال للتمييز التعبير أليلية، تم فحص تعدد الأشكال في المنطقة ترميز
GABRB3، GABRA5 وGABRG3 للتغاير في عينات من الحمض النووي الجيني من القشرة الدماغية الإنسان. تم تحديد المورثات التي أليل معين انزيم الهضم تقييد المنتجات PCR أو عن طريق التسلسل المباشر. وكانت الترددات أليل تحت سيطرة واضطراب طيف التوحد العينات التي تم تحليلها مقارنة بالقيم التي أعلن عنها في قاعدة البيانات SNP
(http://www.ncbi.nlm.nih.gov) لrs2912582، rs140679 وrs140682 (الجدول
1). وكان تردد البيانات أليلية غير متوفر لrs20318 SNP داخل
GABRB3، ومع ذلك تغاير لهذا SNP كان من الشائع في إطار دراسة السكان (الجدول
1).
الجدول 1. ملخص الترددات أليل لتعدد الأشكال المستخدمة في تحليل التعبير أليلية
تم تحديد التعبير أليلية في عينات متخالف بواسطة RT-PCR، تليها أليل معين تقييد الهضم أو كدنا] تسلسل (الشكل
1). كل 21 الضوابط متخالف تحليلها أظهرت التعبير biallelic المساواة لكل GABA A بالمعلومات SNP جين مستقبلات الوحيدات (الجدول
2 وبيانات تمثيلية في الشكل
1). بالإضافة إلى ذلك، كما أظهرت العينة الضابطة واحد التعبير biallelic المتساوي للrs2912582
GABRB3 3'-UTR SNP (لا تظهر البيانات). حلل أظهرت معظم RTT (أربع من خمس سنوات) وأربعة من ثمانية عينات التوحد الدماغ التعبير biallelic المتساوي للGABA A جينات مستقبلات الوحيدات، ولكن أربعة من ثمانية التوحد بالمعلومات واحد من أصل خمسة عينات RTT بالمعلومات أظهرت التعبير monoallelic أو منحرفا للغاية، تلخيصها في الجدول
3 مع بيانات تمثيلية هو مبين في الشكل
1. كان التعبير عن GABA A مستقبلات النصوص لا يمكن كشفها في عدة RTT والتوحد عينات (ND، الجدول
3)، على الأرجح بسبب مستويات منخفضة جدا من التعبير، التي سبق وصفها
لGABRB3 (13). تم التحقق من جودة النص بواسطة RT-PCR من الجينات التدبير المنزلي،
GAPDH. جميع العينات، بما في ذلك RTT 1748، RTT 1815 وAUT 5173 أظهرت مستويات طبيعية من
GAPDH بواسطة RT-PCR (لا تظهر البيانات)، مما يدل على أن مستويات GABA A مستقبلات نص لا يمكن اكتشافها على وجه التحديد في هذه العينات. العدد الكلي للعينات السيطرة تحليلها
(21 كان) أكبر من عدد RTT
(5) أو التوحد
(8) العينات التي تم تحليلها لكل الجينات، مما يشير إلى أن التعبير monoallelic من 15q11-13 GABA A جينات مستقبلة ليس متغير في عموم السكان ولكن محددة إلى RTT وعينات من مرض التوحد.
 الشكل 1.
الشكل 1. يطبع تحليل 15q11-13 GABA A جينات مستقبلة. (A) الاستشرابية التسلسل تثبت التعبير biallelic متساو من GABRB3 (SNP rs20318) وGABRG3 في عينات السيطرة. تسلسل في لوحات العليا هي الحمض النووي ومتواليات في لوحات أقل الجيني هم من كدنا]. السهام السوداء تبرز SNP في كل اللوني. AUT 797 يظهر واضحا التعبير monoallelic من GABRB3 وGABRG3 وRTT 5214 قد انحرفت التعبير عن أليل T في GABRG3. والتسلسل كلا الاتجاهين الأمامي والخلفي لكل فرد وكانت الاستشرابية ثابت. (B) أليل محددة تحليل تقييد هضم GABRA5 يوضح العادي التعبير المساواة biallelic في الضوابط ومعظم التوحد والعينات RTT. تحتوي لوحة العليا لجنة التنسيق الإدارية I-هضم الحمض النووي الجيني من كل فرد ويحتوي على لوحة أقل المنتجات [كدنا لجنة التنسيق الإدارية I هضمها. لجنة التنسيق الإدارية I يقطع أليل C ولكن لا قطع أليل T من GABRA5 SNP. الضوابط والعديد من التوحد والعينات RTT تثبت التعبير biallelic على قدم المساواة، تم الكشف عن ولكن فقط أليل C من GABRA5 في AUT 5173.
الجدول 2. النمط الجيني وأليلية التعبير النتائج للعينات سيطرة الدماغ
الجدول 3. النمط الجيني والتعبير أليلية النتائج للعينات الدماغ RTT وAUT
سابقا، وجدنا انخفاض كبير في البروتين GABRB3 في مجموعة فرعية من عينات التوحد الدماغ
(13). لتحديد ما إذا كانت الخسارة من وbiallelic (لوب) تعبيرا عن GABA A جينات مستقبلات الوحيدات في عينات التوحد يرتبط مع انخفاض مستويات البروتين GABRB3، قارنا القيم البروتين السابقة GABRB3 تطبيع يحددها طخة مناعية شبه الكمي
(13) بين مجموعات من الأفراد بناء في حالة التعبير أليلية. عينات التوحد مع لوب تعبير عن واحد أو أكثر 15q11-13 GABA A مستقبلات الوحيدات الجينات قد خفضت بشكل ملحوظ للغاية نسبة البروتين GABRB3 مقارنة مع الضوابط والتعبير عن biallelically عينات التوحد (الشكل
2 A). لم يكن هناك اختلاف في البروتين GABRB3 بين عينات الرقابة وعينات التوحد مع GABA biallelic تعبير مستقبلات (الشكل
2 A). وتشير هذه النتائج إلى أن الوضع أليلية التعبير عن 15q11-13 GABA A مستقبلات الجينات الوحيدات تتوقع العيوب البروتين التعبير عن GABRB3 في التوحد.
 الرقم 2.
الرقم 2. GABRB3 البروتين ونص المستويات في عينات التوحد الدماغ.
(A) مستويات البروتين GABRB3 المنشورة سابقا
(13) كنسبة يعني ± SEM من جميع العينات في كل فئة وتمت مقارنة بين الضوابط (شريط أبيض)، وعينات من التوحد مع التعبير biallelic (شريط رمادي فاتح) وعينات من التوحد مع لوب 15q11-13 GABA تعبير مستقبلات واحد على الأقل الجينات جبر (لوب، شريط رمادي غامق). عدد الأفراد المدرجة في كل فئة يظهر بين قوسين إلى جانب تسمية كل
خ -axis. عينات لوب التوحد أظهرت أقل بكثير تعبير البروتين GABRB3 من الضوابط
(P = 0.0004) وعينات التوحد biallelic
(P = 0.004)، ولكن لم يكن هناك اختلاف كبير (NS) بين الضوابط وعينات التوحد biallelic. تم قياس
(B) مستويات النص
GABRB3 في التوحد والسيطرة على عينات القشرة المخية التي كتبها الكمي في الوقت الحقيقي وأظهرت مستويات RT-PCR.
GABRB3 نسخة طبيعية
لGAPDH لعناصر (شريط أبيض، تمثل متوسط ± SEM من سبع عينات) وعينات التوحد الفردية تمثل يعني ± SEM من مكررات ثلاثة لكل عينة التوحد. يشار إلى عينات لوب التوحد مع أشرطة رمادية داكنة والحانات رمادي فاتح تمثل عينات التوحد مع التعبير biallelic. اثنين من أربع عينات لوب التوحد، AUT 1174 وAUT 5173، عرضت بشكل ملحوظ انخفضت مستويات النص مقارنة مع الضوابط.
الكمي في الوقت الحقيقي RT-PCR ثم يقوم بنفس عينات [كدنا التوحد الدماغ تحليلها في الجدولين
2 و
3 لتحديد ما إذا كان الوضع التعبير أليلية يعكس مستوى النص الكامل. وعلى النقيض من اختلافات واضحة في بروتين GABRB3، كانت مستويات النص
GABRB3 المتغيرة في عينات التوحد لوب التعبير. شخصين (AUT 1174 وAUT 5173) أظهرت انخفاضا كبيرا
GABRB3 نسخة وشخصين (AUT 797 وAUT 4925) أظهرت مستويات النص لا تختلف كثيرا من الضوابط (الشكل
2 B). تم تنفيذ الكمية RT-PCR أيضا
لGABRG3 وGABRA5. وكانت مستويات التعبير عن
GABRG3 وGABRA5 على ما يبدو 1000 أضعاف أقل من
GABRB3 عندما تطبيع
GAPDH، ولكن الضوابط والتوحد والعينات RTT أظهرت انخفاض مستويات مماثلة من التعبير عن كل من
GABRG3 وGABRA5 (التكميلي المواد، الشكل S1).
لمزيد من التحقيق يطبع الآثار على
GABRB3 التعبير، تم تنفيذ الكمي في الوقت الحقيقي تحليل RT-PCR على PWS وAS [كدنا الدماغ مع الحذف 15q11-13 الأب أو الأم، على التوالي (الشكل
3 A). كانت تسيطر الاختلافات في نوعية الحمض النووي الريبي لتطبيع
GAPDH الجينات التدبير
المنزلي. كان
GABRB3 نسخة أقل بكثير من عينات مراقبة في كل PWS وAS عينات الحذف، ولكن كان عينات حذف PWS التعبير أقل بكثير من عينات AS الحذف، مما يشير إلى وجود تحيز الأب في التعبير الجيني عند منطقة 15q11-13 واحدة موجودة. PWS عينات UPD الأمهات، على الرغم من وجود نسختين من
GABRB3، عرضت انخفاضا كبيرا التعبير
GABRB3 بالمقارنة مع الضوابط، مما يشير إلى وجود تحيز التعبير الأب في حالة عدم وجود متحدر من والدين كروموسوم 15 مساهمة.
 الرقم 3.
الرقم 3. الكمية RT-PCR من 15q11-13 الجينات في القشرة الإنسان. تم إجراء الكمي التضخيم RT-PCR من GABRB3، UBE3A وSNRPN في التجارب ثلاث نسخ على بعد الوفاة كدنا] دماغ الإنسان من السيطرة، RTT، AS الحذف، PWS الحذف وPWS عينات UPD الأمهات. يتم عرض عدد من الأفراد تحليل لكل فئة بين قوسين تحت تسمية س -axis. تم تطبيع جميع الجينات ضد GAPDH تحكم الجينات التدبير المنزلي والقيم -axis ص تمثل يعني ± SEM من كل يعيد لجميع الأفراد في كل فئة. و(A) مستويات GABRB3 نسخة انخفاضا كبيرا في جميع الفئات اضطراب مقارنة مع الضوابط (يشار -values P مع العلامات النجمية ومدرجة أدناه الشكل). AS الحذف وعينات حذف PWS تمت مقارنة وكانت أيضا مختلفة إلى حد كبير. (B) UBE3A التعبير كان أقل من ذلك بكثير في RTT وAS عينات الحذف وأعلى بكثير في عينات PWS UPD. (C) SNRPN التعبير كان غائبا في PWS الحذف وعينات UPD وانخفاض كبير في AS عينات الحذف.
UBE3A، جينة مطبوع أعربت أمهات،
وSNRPN، وقد تم تحليل الجينات مطبوع أعرب أبويا كما جينات التحكم لالكمي RT-PCR مقايسة (الشكل
3 B و C). وعينات حذف AS أظهرت انخفاضا كبيرا التعبير
UBE3A، أظهرت كما هو متوقع بالمقارنة مع الضوابط العينات حذف PWS أي تغيير في
UBE3A التعبير، وكان PWS عينات UPD الأمهات ارتفاع كبير في مستويات
UBE3A (الشكل
3 B). كما هو متوقع، كان
SNRPN غائبة في PWS الحذف وعينات UPD (الشكل
3 C). AS عينات الحذف، ومع ذلك، أظهرت وجود خلل خفية لكنه مهم من المستغرب في
SNRPN (الشكل
3 C).
وقد أظهرت عينات مخ الفأر -deficient كلا RTT
وMecp2 سابقا من نقص في البروتين كبيرة GABRB3
(13). كما عرضت
Mecp2 الماوس -deficient الدماغ عيب النسخي في
Gabrb3، ولذلك أدرجت عينات الدماغ RTT في التحليل
GABRB3 نسخة الكمي. كما هو متوقع من الدراسات الماوس، لوحظ وجود خلل كبير في
GABRB3 نص في RTT المخ بالمقارنة مع التحكم (الشكل
3 A). على الرغم من أن تم الإبلاغ عن بيانات متضاربة حول التقلبات من
Ube3a في
Mecp2 الماوس -deficient
(13، 31، 32)، وقد وجد انخفاض ملحوظ في
UBE3A التعبير في RTT الدماغ (الشكل
3 B)، بما يتفق مع البروتين انخفضت لوحظ سابقا (
13). على الرغم من MeCP2 بربط المروج من
SNRPN (13، 26)، لم يلاحظ أي فرق في
SNRPN نص (الشكل
3، بما يتفق مع صيانة يطبع العادية
للSNRPN في RTT (C)
33)، والتعبير العادي في
Mecp2 - الدماغ نقص الماوس
(13، 34). العيوب
GABRB3 التعبير في RTT
وMecp2 دماغ الفأر -deficient
(13) وقد تورط MeCP2 في تنظيم جينية من 15q11-13 GABA A مستقبلات. بالإضافة إلى ذلك، لوب التعبير عن 15q11-13 GABA A المستقبلات في عينة RTT واحدة وعينات التوحد متعددة مع وصف سابقا MeCP2 عيوب التعبير
(30) تشير إلى أن MeCP2 يلعب دورا في تنظيم هذه الجينات. لتحديد ما إذا MeCP2 تشارك مباشرة في تنظيم GABA 15q11-13 A مستقبلات الجينات الوحيدات، لونين مناعي (رقاقة) قد أنجز على لونين عزل من خلايا العصبية SH-SH5Y الإنسان سلطة النقد الفلسطينية متباينة. وقد سبق أن أظهرت MeCP2 لربط المواقع الدليل السياسي الشامل مميثل بزخارف المجاورة تسلسل A / T
(35)، لذلك تم تصميم الاشعال الشذرة بناء على هذه المعايير. وقد تبين المروج الميثيلي وأمهات من
SNRPN أن يكون الهدف من MeCP2، وكان يستخدم كعنصر تحكم إيجابية
(13، 26). وقد تم تحديد منطقتين داخل 5 'نهاية
GABRB3 ومنطقة واحدة ضمن 5 "نهاية
GABRG3 إيجابية بالنسبة للMeCP2 ملزمة الشذرة (الشكل
4 A). مواقع ملزمة رقاقة إيجابي ضمن المناطق intronic نحو 5 "نهاية الجينات. التجليد كان MeCP2 لا يمكن كشفها في موقع مماثل قرب نهاية 5 'من
GABRA5.
 الرقم 4.
الرقم 4. MeCP2 بربط المواقع الدليل السياسي الشامل مثيلة داخل سلاسل intron من
GABRB3 وGABRG3. (A) لونين مناعي من 48 ساعة التفريق بين الخلايا SH-SY5Y. DNA يمثل مساهمة لونين قبل مناعي، MeCP2 أب يمثل ونين immunoprecipitated مع MeCP2 الضد-C المحطة، وقبل المناعي (سلبي) مراقبة الأجسام المضادة. وتظهر الجينات يعاير للربط إلى يسار اللوحة. وقد استخدم المروج
SNRPN كعنصر تحكم إيجابية
(26). تظهر منطقتين من
GABRB3 ومنطقة intronic من
GABRG3 كشف MeCP2 ملزمة، ومع ذلك، MeCP2 ملزمة
لGABRA5 كان لا يمكن كشفها.
(B) تخطيطي من 2.5 كيلوبايت من 5 'نهاية
GABRB3. يشار إلى الإكسونات الترميز مع مربعات مرقمة سوداء وصناديق رمادية تمثل 5'-UTR. خط متقطع فوق التخطيطي الجين يمثل موقع الجزيرة الدليل السياسي الشامل "نهاية 5. خطوط سوداء مع السهام أقل من الجينات تظهر الوضع النسبي للمنتجات الشذرة PCR (A) وخطوط مع الدوائر المرقمة أعلاه تظهر الجينات الوضع النسبي للمنتجات التسلسل بيسلفيت. وأظهرت نتائج التسلسل بيسلفيت من 48 ساعة متباينة الخلايا SH-SY5Y أدناه التخطيطي الجينات. كل سطر يمثل استنساخ الأفراد والدوائر وتشير المواقع الدليل السياسي الشامل. الدوائر مليئة تمثل المواقع الدليل السياسي الشامل للمثيلة ودوائر شاغرة تمثل المواقع الدليل السياسي الشامل unmethylated. النجوم فوق مواقع مثيلة بينت المواقع الدليل السياسي الشامل بزخارف تسلسل A / T المجاورة التي هي مواقع الربط قابلية عالية MeCP2
(35). (C) تخطيطي من 2.0 كيلوبايت من 5 'نهاية
GABRG3 تبين تسلسل intronic ميثليته للغاية، كما هو موضح أن تكون إيجابية لMeCP2 ملزمة في (A).
للتأكد من أن النتائج الإيجابية الشذرة كان من المقرر أن ربط MeCP2 إلى المواقع الدليل السياسي الشامل ميثليته، تم إجراء التسلسل بيسلفيت. كان الحمض النووي الجيني من الخلايا SH-SH5Y بيسلفيت تحويلها وصممت الاشعال داخل المناطق رقاقة إيجابي عندما يكون ذلك ممكنا. الرقم
4 B و C توضح مناطق
GABRB3 وGABRG3 التي تم تحليلها. على الرغم من أن المناطق تحليلها من الجزيرة
GABRB3 الدليل السياسي الشامل كانت unmethylated تماما تقريبا، ومنطقة محددة MeCP2 المجاورة إنترون 3 أظهرت مثيلة الجزئي (الشكل
4 B). وبالمثل، التسلسل بيسلفيت المنطقة متجهة MeCP2 من
GABRG3 أظهرت مستويات عالية من الدليل السياسي الشامل مثيلة (الشكل
4 C) كذلك يشير إلى أن كل من هذه السلاسل الجينومية وأهداف MeCP2.
تم تحليل الحمض النووي الجيني الدماغ البشري من خلال التسلسل بيسلفيت لتحديد ما إذا كان مثيلة لوحظ في الخلايا SH-SY5Y موجود أيضا في الدماغ. وقد تم تحليل 10 على الأقل الحيوانات المستنسخة لكل فرد وعدد من الأفراد المدرجة في كل فئة والمدرجة أسفل كل رسم بياني (الشكل
5). النسبة المئوية للمواقع مثيلة تعكس عدد من المواقع الدليل السياسي الشامل مميثل مقسوما على جميع المواقع مثيلة المحتملة التي تم تحليلها. عينات الدماغ السيطرة أظهرت أنماط مثيلة مماثلة لمثيلة لوحظ في الخلايا SH-SY5Y (الشكل
4 B و C)، كما هو موضح في الشكل رقم
5 أ و ب.
 الرقم 5.
الرقم 5. التسلسل بيسلفيت من الدماغ البشري والحمض النووي الدم. تم تنفيذ التسلسل بيسلفيت على الحمض النووي الجيني البشري من السيطرة، PWS، AS والأفراد التوحد. وقد تم تحليل 10 على الأقل الحيوانات المستنسخة لكل فرد المدرجة، ويعكس في المئة مثيلة نسبة المواقع الدليل السياسي الشامل مثيلة من العدد الإجمالي للمواقع الدليل السياسي الشامل يعاير لجميع الحيوانات المستنسخة من كل عينة. يتم رسم النتائج على النحو يعني ± SEM من مثيلة في المئة لجميع العينات المدرجة في كل فئة. يتم عرض عدد من الأفراد تحليلها في كل فئة تحت تسمية
س -axis.
(A) GABRB3 تسلسل intronic ميثليته هو مبين في الشكل
4 كان (B) بيسلفيت التسلسل في الدماغ الحمض النووي الجيني من السيطرة، PWS الذكور، PWS أنثى، AS وعينات من مرض التوحد. لم PWS عينات الذكور وعينات التوحد ليس لديها مثيلة تختلف كثيرا من الضوابط. في المقابل، فإن الأنثى عينات PWS وAS العينات أظهرت انخفاضا كبيرا مثيلة
GABRB3. (B) تسلسل ميثليته من
GABRG3 إنترون 1 هو مبين في الشكل
4 كان (C) بيسلفيت التسلسل في السيطرة، PWS، AS والتوحد عينات الدماغ البشري. جميع العينات التي تم تحليلها أظهرت مماثل مستوى عال من مثيلة. ومقارنة
(C) التحكم في الدماغ الحامض النووي من
GABRB3 إنترون 3 مع مثيلة في عينات الدم سيطرة الإنسان وعينة من الحيوانات المنوية واحد. عينات الدم والسائل المنوي انخفاضا في مثيلة مقارنة مع الدماغ
(P <0.02 في الدم). وتمت مقارنة
(D) التحكم في الدماغ ومثيلة الدم المستويات
لGABRG3 إنترون 1 و لم يلاحظ أي اختلاف كبير.
لتحديد ما إذا كان مثيلة لاحظ الوالدين، PWS الحذف، AS الحذف وPWS الأم عينات UPD تم تحليل أليل معين. الخلافات البارزة ومن المثير للاهتمام، وعينات PWS معروضة في
GABRB3 مثيلة على أساس الجنس، لم تراع الفوارق بين الجنسين ولكن مشابهة لعناصر أو فئات اضطرابات أخرى. كل من الإناث PWS وAS عينات (الذكور والإناث) أظهرت أوجه قصور مهمة في مثيلة
لGABRB3 إنترون 3 بالمقارنة مع عينات التحكم (الشكل
5 أ)، مما يوحي بأن مثيلة
GABRB3 ليس بين الوالدين ومن أصل معين ولكن قد تعتمد على وضعها الطبيعي مساهمة الكروموسومات متحدر من والدين. وعلى النقيض من
GABRB3، لم استنساخ
GABRG3 لا تظهر فروق ذات دلالة إحصائية في مثيلة بين الضوابط وPWS / AS العينات، على الرغم من أن المستويات العامة للمثيلة كانت عالية (الشكل
5 B). وقد تم تحليل عينات التوحد الدماغ للتحقيق في احتمال تشوهات الحامض المساهمة في التعبير لوب في بعض هذه العينات. لم يلاحظ أي فرق في مثيلة بين العينات التوحد وضوابط
لGABRB3 أو
GABRG3 (الشكل
5 أ و ب).
لاختبار الفرضية القائلة بأن مثيلة intronic من
GABRB3 يرتبط بشكل إيجابي مع التعبير، ونحن ننظر في أنماط مثيلة في مصادر الأنسجة الإضافية. تحليل
GABRB3 مثيلة في الدم والسائل المنوي وكشف مستوى أقل بكثير من مثيلة مقارنة مع القشرة (الشكل
5 C)، مما يعني أن زيادة مثيلة من
GABRB3 إنترون 3 تكتسب biallelically في الدماغ. وعلى النقيض من الأنسجة محددة
GABRB3 مثيلة، كان
GABRG3 مثيلة عالية على قدم المساواة في الدماغ والدم (الشكل
5 D).
لأن الكروموسوم البشري 15q11-13 متورط في الاضطرابات العصبية النمائية متعددة، فمن الأهمية بمكان أن نفهم كيف يتم تنظيم الجينات في هذا الموضع والتعبير عنها. في هذه الدراسة، لدينا حل النزاع حول يطبع من 15q11-13 GABA كتلة المستقبل من خلال إظهار أن هذه الجينات يتم التعبير عن biallelically في القشرة الجبهية الإنسان حتى الآن يتم التعبير عن aberrantly في الاضطرابات العصبية النمائية متعددة. كما توفر البيانات لدينا أدلة على التأثيرات
العابرة مثلي على مستويات التعبير عن
GABRB3، كعينات مع الوالدين إظهار مساهمة التعبير دون المستوى الأمثل واحد. حددنا كل من الآثار
العابرة والجنس على مثيلة الدماغ محددة من تسلسل intronic من
GABRB3 التي هي بمثابة موقع ملزم للMeCP2. وأخيرا، قدمنا الأدلة الداعمة التي MeCP2 غير منظم الإيجابي
للGABRB3 التعبير واقتراح دور الآلية رواية لMeCP2 كمنشط في التعبير الجيني.
بينما يطبع يتميز جيدا لبعض 15q11-13 الجينات، مثل
SNRPN (36) وUBE3A (27، 28)، ووضع يطبع من GABA قد شكك كتلة مستقبلات
(+18 -
21). قمنا بتحليل مباشرة التعبير أليلية من 15q11-13 GABA A مستقبلات الجينات الوحيدات في الدماغ البشري وأفيد بأن
GABRB3، GABRA5 وGABRG3 يتم عادة أعرب biallelically في القشرة المخية الجبهية، ودعم النتائج السابقة في الماوس
(14، 15). ومن المثير للاهتمام، اكتشفنا وجود تحيز الأب تعبيرا عن
GABRB3 في عينات الدماغ مع واحدة 15q11-13 المساهمات الأبوية، كما لوحظ سابقا في PWS الإنسان وخطوط الخلايا AS lymphoblastoid
(18، 19) وبوساطة microcell نقل كروموسوم
(17). نحن نفترض أن يتم التعبير عن
GABRB3 أكثر كفاءة من كروموسوم الأب من كروموسوم الأمهات في عزلة لأن كروموسوم الأب، تحتوي على العديد من الجينات مطبوع أعرب أبويا، لديها بطبيعتها تكوين لونين أكثر نشاطا من الكروموسوم الأمهات. لم يكن هناك أي دليل على وجود تحيز الأب في سبع عينات مختلفة تحكم متخالف مع المساواة التعبير
GABRB3 biallelic (الجدول
2)، مما يشير إلى أن الأمهات عيب
GABRB3 التعبير انقاذهم من قبل تكامل مع الأب 15q11-13 أليل.
على الرغم من وجود 15q11-13 المساهمات متحدر من والدين، RTT واحدة وأربع عينات التوحد الدماغ أظهرت خسارة التعبير biallelic متساو من واحد أو أكثر من GABA 15q11-13 A جينات مستقبلات الوحيدات (الجدول
3). على الرغم من أن لوحظ لوب التعبير عن كل ثلاثة من 15q11-13 GABA A مستقبلات الجينات الوحيدات، كان
GABRB3 وأعرب معظم غاية وهي الأكثر ملاءمة لاضطرابات التوحد. عينات التوحد مع لوب التعبير عن أي واحد من GABA 15q11-13 وكان مستقبلات الجينات الوحيدات انخفاض كبير في مستويات البروتين GABRB3 مقارنة العينات مع التعبير biallelic. وعلاوة على ذلك، عينة واحدة، AUT 797، متخالف لجميع GABA A ثلاثة جينات مستقبلة، أظهر لوب التعبير لكلا
GABRB3 وGABRG3 (الشكل
1). هذه الملاحظات تشير إلى أن العيوب في عينات التوحد موجودة في جميع أنحاء GABA 15q11-13 كتلة المستقبل وربما يرجع ذلك إلى التقلبات جينية من أليل واحد الوالدين. وكانت عينات من الحمض النووي الأبوية غير متوفرة لعينات الدماغ بعد الوفاة، وبالتالي فإن أصل الوالدين من أليل أعرب غير معروف حاليا.
كنا نتوقع
GABRB3 نص على أن تخفض بشكل كبير في العينات مع لوب التعبير، ومع ذلك، مستويات النص المترابطة للبروتين انخفاض في اثنين فقط من أربع عينات التوحد مع لوب التعبير. هذا التناقض يمكن تفسير جزئيا خلفيات وراثية معقدة من عينات التوحد. تم الكشف عن أية طفرات ضمن تسلسل ترميز
GABRB3 في عينات مرض التوحد، ولكن الاختلافات في تسلسل التنظيمية قد يؤثر بشكل كبير النسخ والاستقرار النصوص. ومن المثير للاهتمام،
وGABRB3 أفيد تعدد الأشكال المروج مؤخرا لإضعاف النشاط النسخي في مقايسة luciferase المراسل
(37)، ولكن تم العثور على هذا التعدد في كثير من الأحيان ضمن عينات لدينا، ولا يمكن تفسير الاختلافات في
GABRB3 التعبير (لا تظهر البيانات). بدلا من ذلك، الفروق الفردية في التنظيم بعد النسخي من
GABRB3 داخل عينات التوحد قد يفسر الاختلافات بين النص والبروتين. بالإضافة إلى ذلك، GABA A وقد ثبت مستقبلات الوحيدات الجينات لعرض مجمع تنسيق تنظيم
(38)، وبالتالي قد تكون مسؤولة آلية التعويض عن الاختلافات النسخي لوحظ في بعض العينات التوحد الدماغ.
وتمشيا مع الجينات وأعرب بنشاط، والمروج من
GABRB3 كان hypomethylated، ولكن منطقة intronic المجاورة ومميثل أكثر شدة في الدماغ البشري مما كانت عليه في الدم (الشكل
5 C). وتبين لدينا بيانات الوقت الحقيقي RT-PCR الكمية التي القشرة الدماغية لديها مستويات عالية من
GABRB3 التعبير (الشكل
3 أ)، في حين أن الخلايا lymphoblastoid لها بالكاد يمكن كشفها
GABRB3 مستويات التعبير أقل من 1/1000 من مستويات النص الدماغ (لا تظهر البيانات) . الحامض النووي داخل المناطق intronic أو بين الجينات وقد وصفت سابقا لإضفاء تأثير إيجابي على التعبير الجيني (39،
40)، ولذا فإننا نقترح أن مثيلة intronic من GABRB3 يرتبط بشكل إيجابي مع التعبير. في حين PWS وAS عينات الدماغ لم تكشف الأم المنشأ أنماط مثيلة محددة في
GABRB3 وGABRG3، AS عينات الحذف وعينات الإناث PWS خفضت بشكل ملحوظ من مستويات
GABRB3 إنترون مثيلة. وhypomethylation كل من عينات AS وPWS تشير إلى أن
العابرة الآثار بين الأليلات الأبوية تعمل على تنظيم مستويات الحامض المناسبة في هذا الموضع في القشرة. الفرق بين الجنسين في عينات PWS
لGABRB3 كان غير متوقع، ولكن، Liljelund وآخرون. (41)، كما وجدت بين الجنسين وبين الوالدين والمنشأ الآثار على Gabrb3 تعبير البروتين في الفئران متخالف لصغير
Gabrb3 الحذف التي تضم منطقة syntenic للمواقع مثيلة إنترون 3. لم عينات السيطرة والتوحد الدماغ لا يحمل آثار الجنسين على مثيلة والتوحد العينات مع لوب التعبير لم يكن لديهم عيوب مثيلة داخل المناطق يعاير من
GABRB3 أو
GABRG3 (الشكل 5). ربما، وانخفاض كبير في التعبير MeCP2 الملاحظة في عينات التوحد مع التعبير لوب (30،
42 قد) يكون الخلل الجيني الرئيسي مما تسبب dysregulated التعبير GABRB3. بالإضافة إلى ذلك، عيوب مثيلة قد تكون موجودة في المناطق البعيدة داخل أو بالقرب
GABRB3 باستخدام تقنيات عالية الإنتاجية في عينات التوحد مع لوب GABA 15q11-13 A التعبير مستقبلات في الدراسات المستقبلية.
وصفناها سابقا عملية تنظيم تنمويا من الاقتران مثلي من الإنسان 15q11-13
GABRB3 الأليلات في الدماغ الطبيعي أن تعاني من عجز في PWS، AS، RTT وعينات التوحد (26). أوجه القصور في الخلايا العصبية
عبر تم تفاعلات 15q11-13 المجالات ارتباطا مباشرا MeCP2 التي كتبها الاجتثاث من MeCP2 ملزمة في الخلايا العصبية SH-SY5Y أن يخفف كثيرا من الاقتران من
GABRB3 الأليلات (26). على الرغم من MeCP2 طفرة أو نقص ليست كافية للتسبب لوب التعبير عن 15q11-13 GABA A جينات مستقبلات في جميع العينات RTT والتوحد، نتائجنا تشير إلى أن
عبر التفاعلات بين المتماثلات 15q11-13 الوالدين تسهم في السليم العادي التعبير الجيني biallelic داخل GABA A مجال مستقبلات. الانخفاض الكبير غير المتوقع
للSNRPN التعبير في عينات AS الحذف (الشكل
3 C) تشير إلى أن
عبر تفاعلات مهمة للحصول على أفضل مستويات التعبير الجيني داخل 15q11-13. وتشير البيانات المتوفرة لدينا أن التعبير monoallelic من
GABRB3 لا تحدث في الدماغ العادي، مما يوحي بأن التعبير biallelic من
GABRB3 يتم الاحتفاظ بنشاط. نحن نفترض أن قربها من الكتلة مستقبلات GABA 15q11-13 إلى الجينات مطبوع يتطلب عملية نشطة فيه كل
GABRB3 الأليلات المنتسبين فعليا من أجل أن تتجلى على النحو الأمثل من كل الأليلات.
عينات RTT مع التعبير biallelic أيضا خفضت بشكل ملحوظ
GABRB3 التعبير (الشكل
3 أ)، تورط دورا إيجابيا لMeCP2 في تنظيم
GABRB3 التعبير عن طريق الربط بين المواقع الدليل السياسي الشامل مثيلة في كل من
رابطة الدول المستقلة والعابرة. نموذج هو موضح في الشكل
6 يوضح احتمالين لتفسير انخفاض biallelic والتعبير monoallelic من
GABRB3 في RTT الدماغ. في الدماغ العادي، MeCP2 ملزمة لINTRON 3 مواقع مثيلة يعمل على تنظيم إيجابيا
GABRB3 التعبير من خلال جلب كل من الأليلات الأبوية إلى موقع نووي مع الآلات النسخي المثلى، مثل مصنع النسخ (43). في حالات انخفاض التعبير biallelic، لا يتم إقران 15q11-13 المتماثلات بسبب النقص في MeCP2، ونسخه لا كروموسوم الوالدين على النحو الأمثل. وبالمثل،
Mecp2 الفئران -deficient أيضا يحمل انخفاض شقين كبير في
Gabrb3 على الرغم من التعبير biallelic العادي (13). في التوحد والعينات RTT مع التعبير monoallelic، وتعطلت الاقتران، ولكن جينية إضافية أو خلل جيني يمنع النسخ من أليل واحد (الشكل 6). على الرغم من أننا التكهن بأن أليل الأمهات صامت، وذلك بسبب أدلة على وجود تحيز الأب في التعبير (الشكل
3 أ)، وأصل الوالدين من الأليلات في التوحد وRTT عينات بعد الوفاة الدماغ غير معروف حاليا.
 الرقم 6.
الرقم 6. نموذج للمشاركة MeCP2 في التنظيم الإيجابي للGABRB3 التعبير في الدماغ. وتظهر الصبغيات الأبوية للأم وكما الوردي (الأم) والأزرق (الأب) الخطوط المنقطة، على التوالي. حجم السهام يمثل المستويات النسبية في التعبير الجيني. وحدة النسخ الأب محددة كبيرة من SNRPN من خلال نص العقاقير من UBE3A هو مبين في شكل سحابة. في عينات المخ المعتادة MeCP2 بربط المواقع الدليل السياسي الشامل مثيلة داخل إنترون من GABRB3 على كل من الكروموسومات ويعزز GABRB3 التعبير والاقتران مثلي كتبها توطين كل من الأليلات لمساحة النسخ نشط مثل مصنع النسخ. في المرضى الذين يعانون متلازمة ريت مع طفرة MeCP2 أو نقص، وتعطلت الاقتران مثلي من 15q11-13، كما هو موضح سابقا في ثاتشر وآخرون. (26). في RTT الأفراد مع انخفاض التعبير biallelic الأليلات كل من الأم والأب وكتب عند مستويات منخفضة بسبب موقعها النووي دون المستوى الأمثل. الأفراد مع لوب التعبير قد يكون لها عيب جينية إضافية مما يؤدي إلى تخفيض الانتقائي للأليل واحد (ممثلة في X). لأن 15q11-13 الحذف يؤدي إلى فقدان الاقتران مثلي والتحيز الأب في GABRB3 التعبير، ونحن نفترض أن الأليل الأب وأعرب عن تفضيلي في مثل هذه الحالات.
في هذا التقرير نعرض بواسطة رقاقة وتسلسل بيسلفيت أن MeCP2 بربط intronic متواليات الدليل السياسي الشامل مثيلة داخل
GABRB3 في خلايا SH-SH5Y الإنسان متباينة (الشكل 4). على الرغم من أن ينظر إليها عادة على أنها قامع النسخي (44،
45)، وقد تم مؤخرا أظهرت MeCP2 غير مباشر لتنظيم إيجابا على الجينات المستهدفة عامل التغذية العصبية الدماغي (46) والتعبير الجينوم على نطاق التنميط التحليلات أثبتت العديد يحتمل أن تكون أسفل تنظيم الجينات مع MeCP2 نقص
(47 - 49). في نموذجنا (الشكل
6)، نقترح أن ربط MeCP2 إلى مواقع الدليل السياسي الشامل مميثل الدماغ محددة داخل GABRB3 يؤثر إيجابيا التعبير من خلال الترويج
عبر التفاعلات بين المتماثلات وتنظيم إيجابيا لونين في
رابطة الدول المستقلة. Horike وآخرون. (34) سبق وصفها دور للMeCP2 كمنظم لونين حلقات هذا التعبير الصمت الجينات. نحن نفترض أن ملزمة لمواقع متعددة في
رابطة الدول المستقلة في GABA A كتلة المستقبل، MeCP2 يعزز تشكيل هياكل لونين مواتية للنسخ النشط، كما يتضح من قربها من مصنع النسخ النشط في النموذج في الشكل 6.
GABA هو الناقل العصبي المثبط الرئيسي في الدماغ، ويلعب دورا مهما في وظيفة الدماغ. انقطاع جزئي من الخلايا العصبية GABAergic في القشرة من الفئران يسبب نوبات والقلق والسلوك الاجتماعي ضعاف، بما يتفق مع GABA مما يشير إلى عيوب المساهمة في مسببات اضطرابات التوحد (50). على عكس GABRB3، GABRA5 وGABRG3 البروتينات لا توطين في المقام الأول في القشرة المخية
(+51 -
53)، بما يتفق مع البيانات التعبير لدينا (المواد التكميلية، الشكل S1). حذف كل من
Gabra5 وGabrg3 لا يسبب النمط الظاهري واضح في الفئران (54،
55)، ومع ذلك، Gabrb3 الفئران -deficient الظواهر المعرض مثل المضبوطات، وتشوهات النوم والسلوكيات النمطية تتفق مع خصائص AS، RTT والتوحد (55).
التداخل الوراثي والمظهري في RTT والتوحد وAS تشير إلى أن تداخل المسارات الجزيئية وdysregulated في هذه الاضطرابات النمائية العصبية مماثلة. وإن كان نادرا،
MECP2 تم الإبلاغ عن الطفرات في الأفراد المشخصة سريريا مع AS أو التوحد (56،
57)، وMECP2 تم الإبلاغ عن الطفرات جنبا إلى جنب مع إعادة ترتيب في 15q11-13 في الأشخاص الذين يعانون RTT (58). في هذه الدراسة تبين لنا أن
GABRB3، جين 15q11-13، وينظم إيجابيا MeCP2 وبشكل ملحوظ انخفاض في كل من RTT وAS. الملاحظات السريرية للشذوذ EEG نموذجية من AS في المرضى الذين يعانون RTT دعم مشاركة
GABRB3 في الظواهر الصرع المشتركة في هذه الاضطرابات (59، 60). الازدواجية الأمهات من 15q11-13، بما في ذلك
GABRB3 تحدث في نسبة صغيرة من الأفراد المصابين بالتوحد، إلا أنه من غير الواضح كيفية تأثير هذه الازدواجية
GABRB3 التعبير في الدماغ. وعلى الرغم من توقع زيادة عدد النسخ لزيادة التعبير، بياناتنا تظهر معقدة
عبر تنظيم
GABRB3 في الدماغ البشري مما يدل على أن الانحراف عن العادي مساهمة الكروموسومات متحدر من والدين قد يؤثر سلبا
GABRB3 التعبير. كما يوفر دراستنا الحالية دليل على التقلبات جينية معقدة من GABA 15q11-13 A كتلة المستقبل في عينات التوحد مجهول السبب مع العيوب البروتين GABRB3، مما يشير إلى أن المسارات الجزيئية المشتركة وdysregulated في الاضطرابات العصبية النمائية ذات الصلة.
المواد التكميلية متوفرة في HMG على الانترنت.
 « آخــر المشـاركــات »
« آخــر المشـاركــات »
 11-20-2015, 08:46 PM
11-20-2015, 08:46 PM










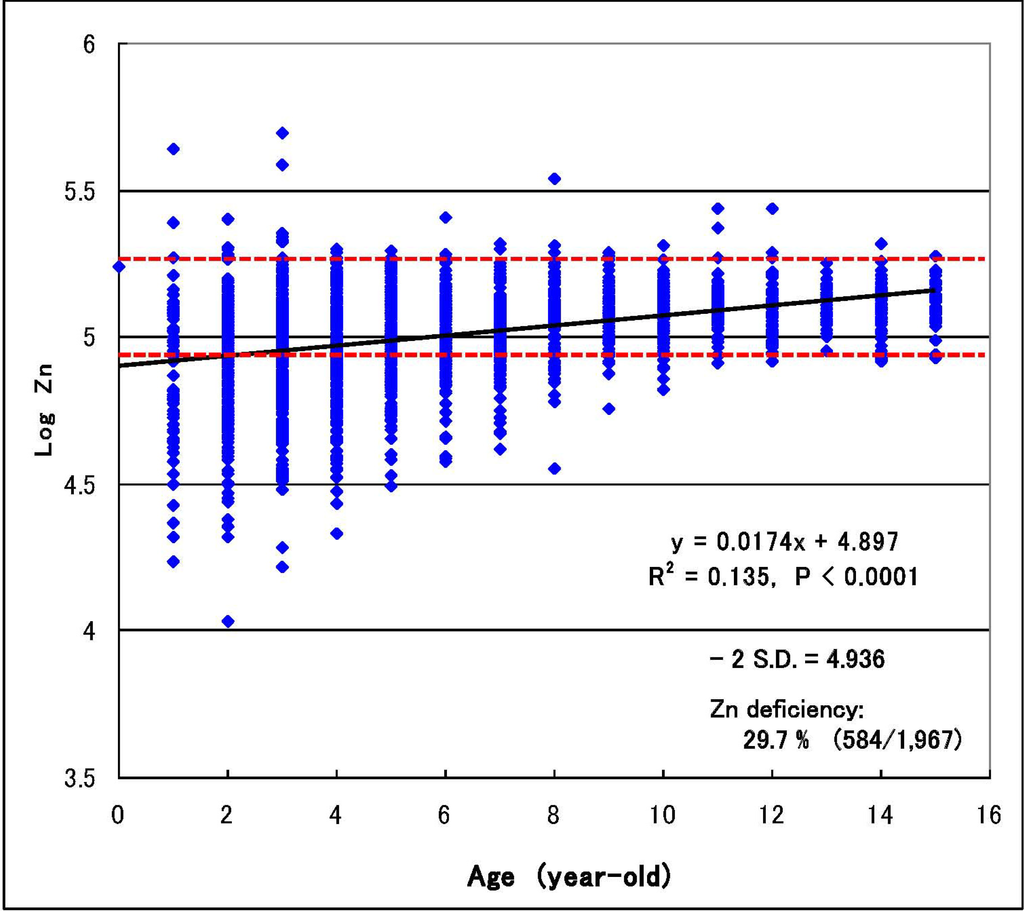
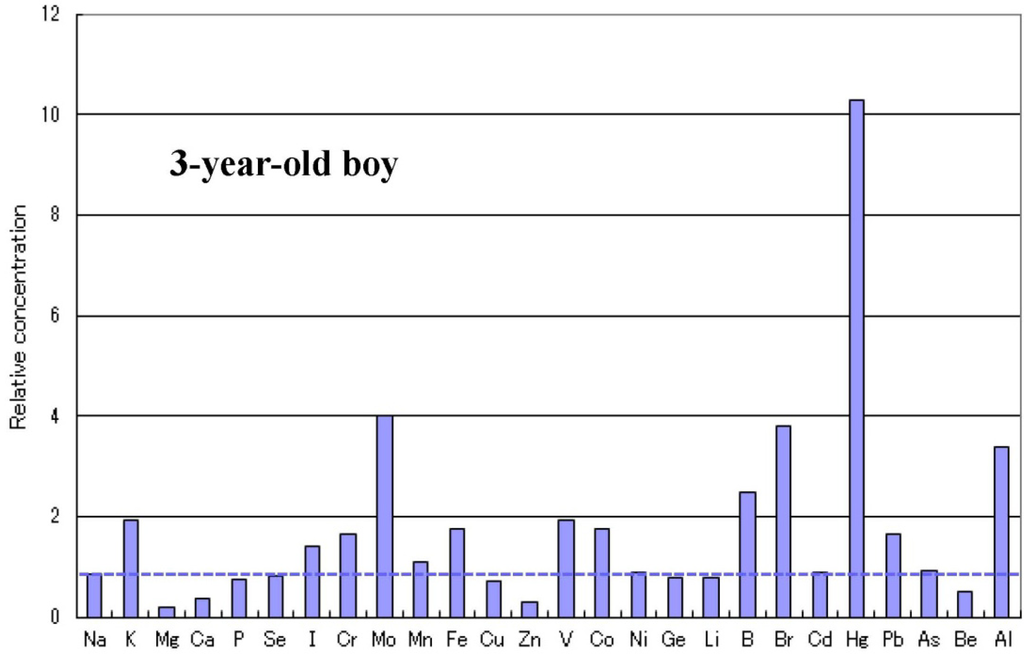
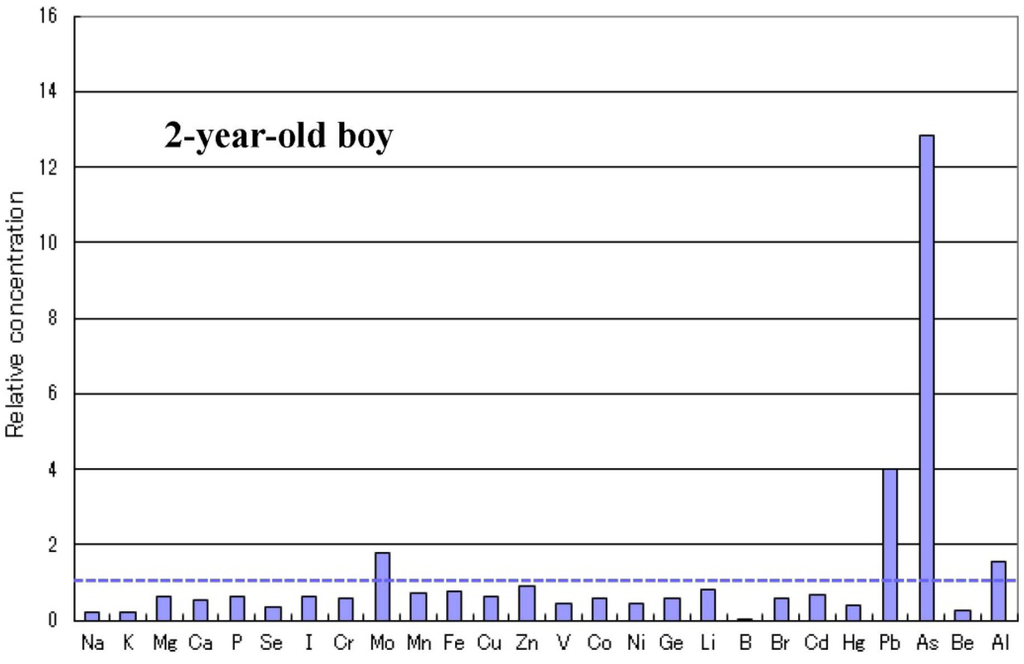
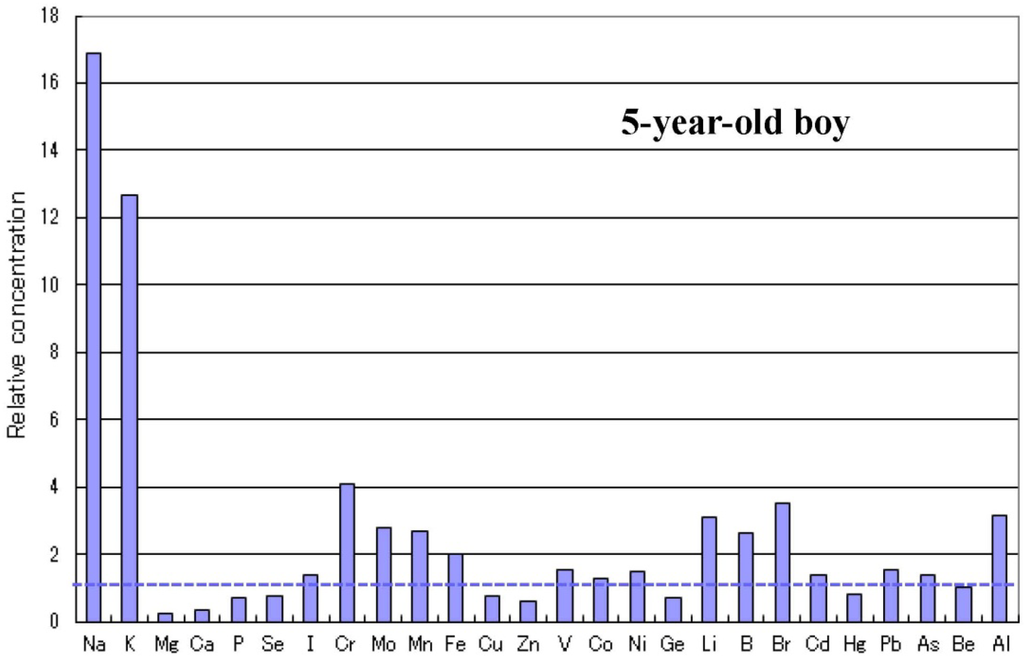
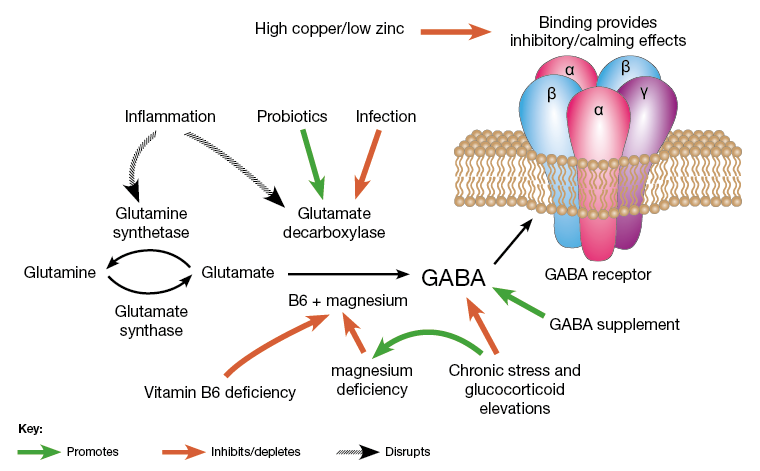
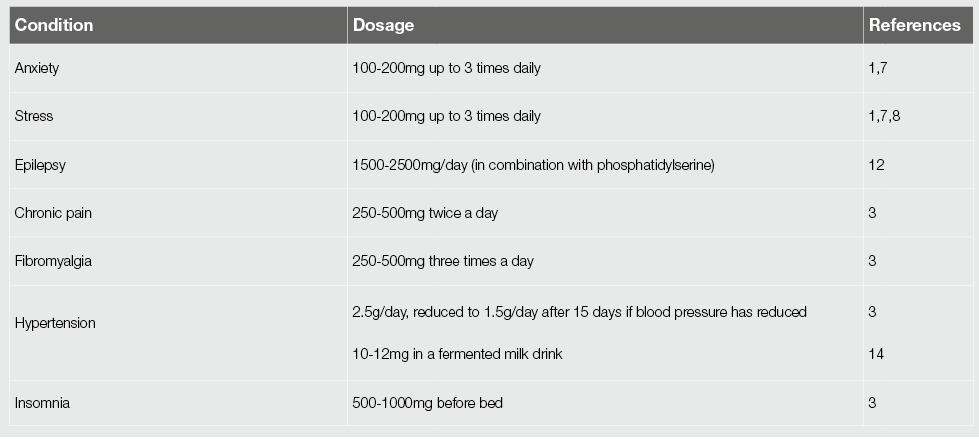
 الجدول 1. انتشار نقص المعادن في الأطفال المصابين بالتوحد
الجدول 1. انتشار نقص المعادن في الأطفال المصابين بالتوحد 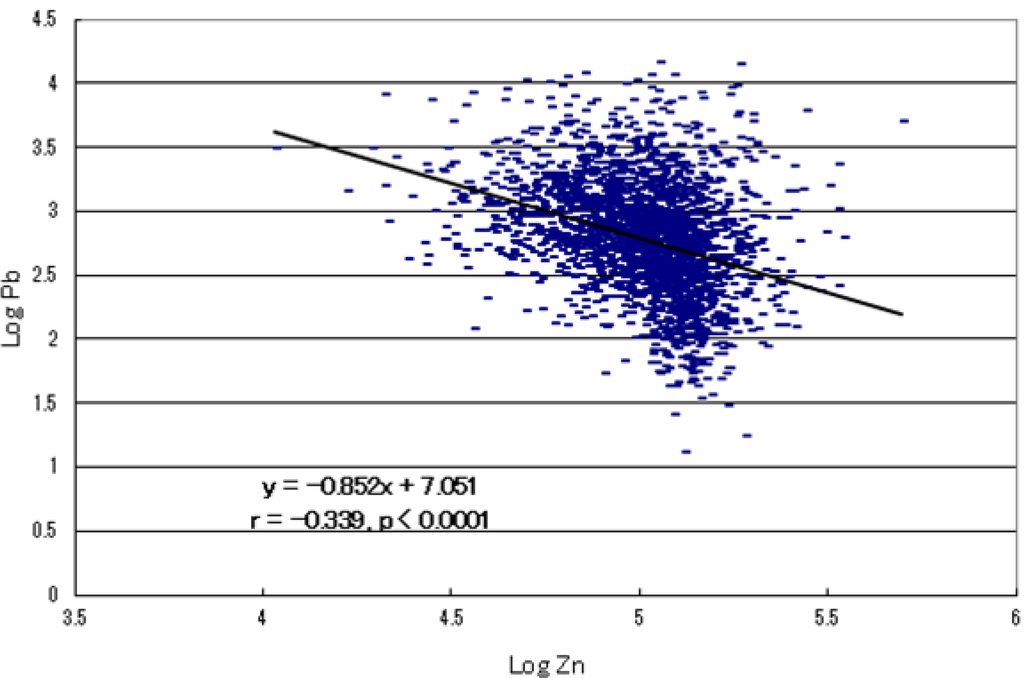
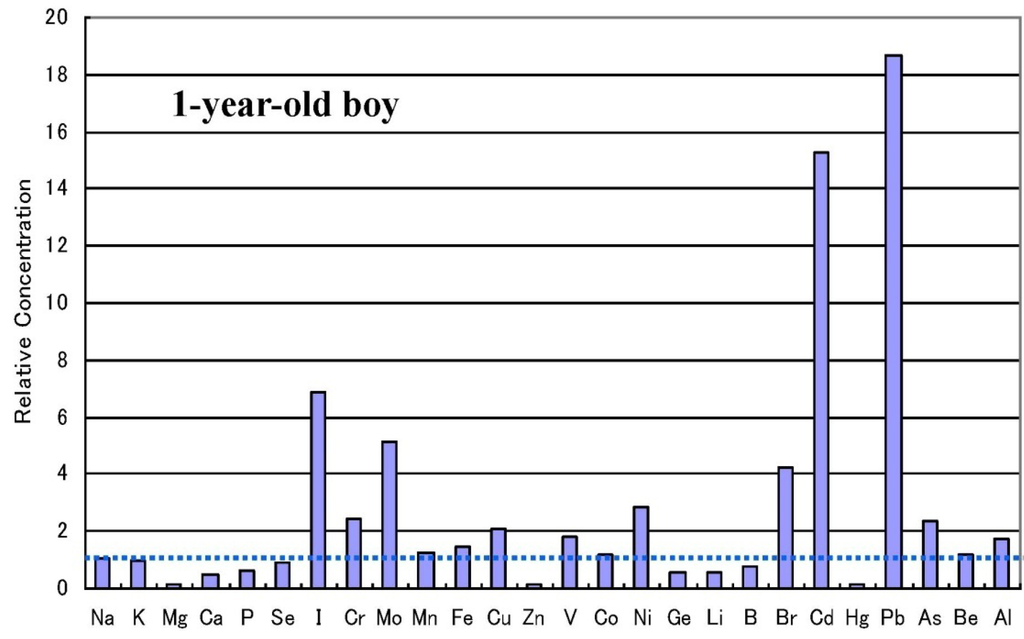
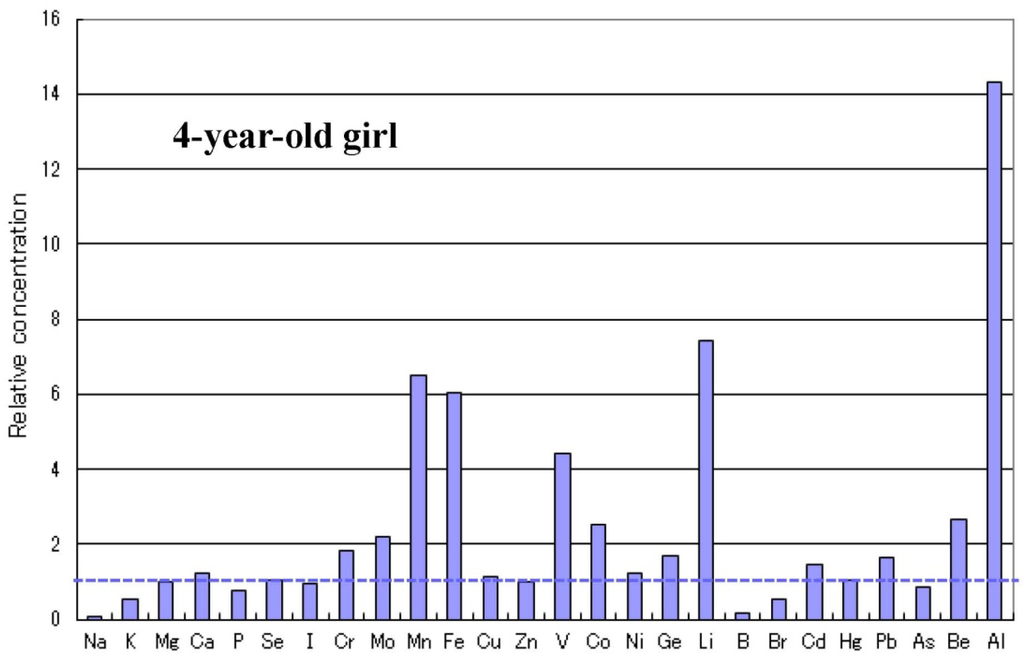












 العرض العادي
العرض العادي

